
1. INTRODUCTION
Artisanal and small-scale gold mining (ASGM) refers to the extraction of gold by local communities or small groups of individuals using traditional methods. This activity is critical to the local economy, particularly in areas with abundant mineral resources. This activity resulted in positive outcomes, such as increased employment and economic growth in the region. This enterprise has grown into a key economic source for hundreds of tribes across Indonesia, spanning regions from Aceh to Papua. ASGM accounts for 76% of miners in West Lombok [1], increases local people’s income by 400–1800% in Sumbawa Island, and creates job opportunities [2].
Although ASGM contributes to local livelihoods, it also causes serious environmental degradation. Gold extraction in ASGM commonly involves ore crushing, tailing deposition, and, in many cases, amalgamation with mercury, all of which can alter soil properties and introduce contaminants into surrounding environments [3]. As a result, former ASGM sites are often characterized not only by mercury contamination, but also by enrichment of other metals, such as arsenic (As), cadmium (Cd), lead (Pb), copper (Cu), zinc (Zn), and iron (Fe) [4], together with soil acidification [5], nutrient depletion, reduced organic matter, and impaired microbial activity [6]. These combined effects reduce soil fertility and limit the capacity of post-mining land to support plant establishment and safe agricultural reuse.
The impact of ASGM is reported to have changed soil pH, nutrient availability, and soil organic content, and to have a direct effect on soil fertility [7]. Excessive mercury concentration in post-ASGM soils altered soil chemical characteristics, including soil fertility [8], decreasing total nitrogen (N) and available phosphorus (P) [9]. Mining activity in ASGM decreased soil pH, inhibiting the microbial community [10], thereby increasing the bioavailability of heavy metals [11]. As, Cd, mercury (Hg), Pb, Cu, and Zn were found to be high in post-mining ASGM [12]. A large release of mercury into the environment may enter the food chain, leading to mercury accumulation in living organisms and thereby increasing the risk to human health. Furthermore, the transformation of inorganic mercury into its more poisonous organic form, methylmercury, markedly exacerbates the environmental and health risks [13,14]. Recognizing this potential impact of ASGM activity, a remediation effort is essential.
Remediation of ASGM-degraded soil is often difficult and costly because contamination occurs alongside broader soil quality deterioration. The complexity of metal mixtures, their variable bioavailability, and the poor physicochemical condition of post-mining soil all increase the technical difficulty of remediation [15]. Conventional approaches, such as excavation, soil covering, thermal desorption, soil washing, electrokinetic remediation, and vitrification are frequently expensive, technically demanding, and difficult to apply at the community scale [16,17]. Therefore, low-cost biological approaches that can simultaneously reduce metal stress and improve soil function are of particular interest for former ASGM land.
Application of a biological agent, such as a selected soil microorganism, may be a promising method, as these organisms have a high capacity to improve heavily degraded soil, such as in mining areas. Plant growth-promoting rhizobacteria (PGPR) and arbuscular mycorrhizal fungi (AMF) are well-known for their mutualistic symbiosis with plants. PGPR contributes to increasing indole-3-acetic acid levels in plants, enhancing heavy-metal chelation in phytoremediation, inducing systemic resistance, and thereby decreasing toxicity [18]. Symbiosis with AMF enhances nutrient uptake, particularly P, in post-mining land [19], and AMF are also recognized for their role as biofertilizers in sustainable agriculture [36], and decreases the translocation factor and bioconcentration of heavy metals (Cd, Pb, Hg) by accumulating them in the rhizosphere or roots through the extra-radicular mycelium of AMF [20,21]. Given their contribution to improving degraded soil, synergistic symbiosis between PGPR and AMF is particularly important for soil remediation in post-gold mining, as they can increase the efficiency of heavy metal remediation and nutrient addition.
Beyond reducing mercury-related risks, the restoration of post-ASGM land should focus on restoring soil functionality as a medium for plant growth. In numerous former ASGM sites, soil degradation is not limited to metal contamination, but also includes low fertility, poor nutrient availability, disrupted microbial activity, and decreased capacity to support crop establishment. Therefore, an effective remediation strategy should not only mitigate contamination, but also improve the biological and agronomic quality of the soil. In this context, microbial-based remediation using consortium AMF+PGPR is particularly relevant, as these microorganisms may simultaneously alleviate heavy metal stress, improve nutrient acquisition, and promote plant growth in degraded soils. However, the specific contribution of PGPR–AMF consortia to soil quality recovery remains difficult to distinguish, since many previous studies have combined microbial inoculation with organic amendments and have mainly focused on tolerant pioneer species rather than sensitive crop plants.
Research on the application of consortium PGPR and AMF to improve heavy-metal-contaminated soil in post-mining environments has been reported, particularly regarding increased plant growth and phytoremediation efficiency. However, most of these studies still focus on relatively tolerant pioneer plants and are generally combined with various organic materials or soil amendments. Unlike previous studies that commonly combine microbial inoculation with organic amendments, this study aimed to evaluate the capacity of a PGPR–AMF consortium alone to restore soil quality in degraded soil from a former artisanal gold mine. The absence of soil amendments in this study was intentionally designed to isolate the biological effect of the AMF–PGPR consortium on soil quality improvement. By maintaining soil as the sole growth medium, the experiment enables direct evaluation of the microbial consortium’s intrinsic capacity to enhance soil fertility and structure, without the confounding influence of external nutrient inputs. This approach provides a clearer understanding of the fundamental role of microbial symbiosis in restoring degraded soil systems.
2. MATERIALS AND METHODS
2.1. Soil Sampling and Soil Preparation
Soil contaminated with ASGM was collected randomly at a depth of 0–30 cm from Kokap District, Kulon Progo Regency, Special Region of Yogyakarta (7°49’37.9”S 110°04’15.6”E and 7°49’38.9”S 110°04’11.2”E). Soil from an undisturbed area of a natural forest (NF) was collected, as well as soil control. Soil was sterilized by steam heating at 100°C for 5 h. Sterilized soil was used to reduce interference from indigenous microbial communities. This step helped improve attribution of observed effects to the introduced AMF–PGPR consortium. The controlled design aimed to test the intrinsic contribution of inoculation under simplified conditions. It was not intended to reproduce the full ecological complexity of field-contaminated soil. However, steam sterilization may alter soil microbial ecology, nutrient dynamics, and potentially metal bioavailability; therefore, the present greenhouse system should be interpreted as a simplified mechanistic model rather than a full representation of natural field conditions. This soil was used as the growth medium for seedlings and is targeted to be treated with PGPR+AMF.
2.2. Seedling Preparation
Seedlings of Capsicum frutescens were prepared by sowing the seeds in sterilized sand. Seedlings were raised by applying only water. One-month-old seedlings were then transferred to the sterilized soil collected in ASGM as a plant indicator. Seedlings of C. frutescens were then put into 500 g of sterilized soil without any additional material or treatment. These seedlings were treated as controls. Other seedlings were received, a consortium of about 20 g of AMF and 20 mL PGPR.
2.3. Inoculum’s Preparation
2.3.1. AMF inoculum preparation
The AMF inoculum used in this research was Rhizophagus clarus (syn. Glomus clarum), obtained from soil at a post-coal mining site in East Kalimantan. AMF spores were extracted using the wet sieving and decanting method followed by sucrose centrifugation. Single-spore isolation was performed under a stereomicroscope. To obtain a monospecific culture, the selected spores were propagated using Sorghum bicolor grown in sterilized zeolite under greenhouse conditions for 3 months. The AMF culture was repeatedly propagated to ensure spore morphological uniformity. The seedlings of S. bicolor were maintained by applying a modified Hoagland solution containing 1 ppm phosphorus (P) once a week. After 3 months, the AMF were harvested, and the AMF spore numbers were counted. There were 90 AMF spores/g. Isolate identification was conducted using molecular approaches [22].
2.3.2. PGPR inoculum preparation
The PGPR inoculum used in this research was collected from post tin mining sites. PGPR used was a consortium of Staphylococcus saprophyticus, Staphylococcus arlettae, and Enterobacter ludwigii. Before inoculation, the bacterial strains were functionally pre-screened for selected plant growth-promoting traits. Two isolates demonstrating phosphate-solubilizing ability were identified by their formation of a clear halo zone on Pikovskaya medium. E. ludwigii was chosen for its nitrogen-fixing potential, as indicated by growth in semi-solid N-free bromothymol blue medium. PGPR inoculum was produced by introducing a single colony of PGPR into 1 L of sterile nutrient broth autoclaved at 121°C, 1 atm for 15 min, and incubating under dark conditions at room temperature (28–30°C) for 72 h. The population of PGPR was 4.8 × 106 colony-forming unit (CFU)/mL prior to inoculation.
2.4. Seedling’s Maintenance
A total of 21 pots (plants) per treatment were used as biological replicates for survival rate measurements. For final seedling’s biomass and laboratory-based analyses (e.g., heavy metal concentration, nutrient content, biomass weight), three plants per treatment were sampled from the 21 replicates due to resource limitations. Pots were randomized in the greenhouse using a completely randomized design, with treatments assigned randomly, and pot positions were rotated weekly to minimize greenhouse position effects. Seedlings were irrigated every 2 days by adding water until the soil reached field capacity. This irrigation regime was applied to maintain stable soil moisture conditions suitable for plant growth while minimizing excessive leaching that could influence nutrient availability and metal mobility. There was no fertilizer applied. Seedlings were grown under greenhouse conditions for 6 weeks without fertilizer application. This experimental duration was considered sufficient to observe early plant establishment and the initial effects of microbial inoculation on soil properties and plant performance.
Seedling’s survival was monitored periodically at 0, 2, 4, and 6 weeks after planting to evaluate plant establishment under each treatment condition. At each observation time, the number of surviving plants in each experimental unit was recorded and compared with the total number of plants initially planted. A plant was classified as surviving when it remained upright and showed visible viable tissues (e.g., green leaves or stem) during the observation period. Plant growth performance was measured at harvest time by assessing the biomass dry weight of both the shoot and the root. AMF colonization was determined as described in Giovannetti and Mosse [23].
2.5. Soil, Shoot, and Root Chemical Analysis
Initial soil samples were collected before planting to determine baseline soil characteristics. Final soil samples, both with and without inoculation, were collected at the end of the experiment (6 weeks after planting) during plant harvesting for analysis of soil chemical properties. Shoots and roots of seedlings were analyzed after harvest. Total metal concentrations (Hg, Fe, Cd, Mn, Zn, and As) in soil, shoot, and root samples were determined using acid digestion with a mixture of nitric acid (HNO3) and hydrogen peroxide (H2O2), following the USEPA Method 3050B [24], and subsequently analyzed using inductively coupled plasma- mass spectrometry. Soil total K was analyzed using the same destruction method, followed by atomic absorption spectrophotometry (AAS) analysis. Soil sulfide was extracted using 0.01 M CaCl2 (1:5 w/v), shaken for 1 h, and filtered. The extract was treated with Buffer A and BaCl2 to precipitate BaSO4. After dissolution of the precipitate according to the laboratory protocol, barium concentration was quantified by atomic absorption spectrometry (AAS), and sulfate concentration was calculated stoichiometrically from the Ba–SO4 molar ratio. The procedure was adapted from the APHA turbidimetric sulfate method (4500-SO42− E). Soil potential phosphorus pentoxide (P2O5) was measured by applying acid-extractable P (hydrochloric acid [HCl] 25%) following the analytical methods described by the Indonesian Soil Research Institute as outlined in Agus et al. [25]. The phosphate fraction in the extract was then determined by the molybdenum blue colorimetric method [26]. Furthermore, soil total nitrogen (N) (Kjeldahl method), soil cation exchange capacity (CEC) (ammonium acetate (1 M, pH 7), soil moisture content (gravimetrically by oven-drying at 105°C for 24 h), soil organic carbon (SOC) (Walkley-Black method), soil pH (H2O) and pH (KCl) were analyzed as well.
2.6. Microbial Analysis
To assess AMF colonization, roots were decontaminated of soil particles, immersed in 70% alcohol overnight, and subjected to 10% potassium hydroxide at 80°C for 15–20 min; thereafter, they were treated with 1% HCl for 5 min. The samples were subsequently stained with 500 mg/L trypan blue. The assessment of AMF colonization was conducted according to the methodology established by Giovannetti and Mosse.
Viable bacterial enumeration was evaluated at the end of the experiment (6 weeks). Rhizosphere soil was collected from the root-surface zone of each polybag (inoculation zone) and enumerated by plate count to assess viable bacterial abundance. CFUs were counted and reported as CFU/mL of soil suspension. All handling was performed using sterile tools to minimize contamination. Viable bacterial counts were determined from three independent replicates for each treatment, and the results were expressed as mean ± standard error (SE) (CFU/mL of soil suspension)
2.7. Statistical Analysis
Statistical analyses were performed using Kaleida Graph version 4.1 (Synergy Software, 2012, USA). Data are presented as mean ± SE of three independent replicates (n = 3). Differences among treatments were evaluated using one-way analysis of variance, followed by Tukey’s honestly significant difference post hoc test at P < 0.05.
3. RESULTS AND DISCUSSION
3.1. Impacts of ASGM on Soil Physicochemical Properties and Nutrient Status
Generally, ASGM markedly decreased the soil physicochemical properties [Figure 1] and soil nutrition status [Figure 2] relative to the non-contaminated reference. Soil pH remained strongly acidic in the contaminated soil. In Figure 1, the non-ASGM soil (NF) had a pH of approximately 5.4, which was consistently the highest, whereas the contaminated soil post mining (PM) remained within the range of 2.8–3.1. Water content, CEC, and SOC, whereas the ASGM-affected media showed substantially lower values. Water content declined from about 8% in NF to 4.5 in the contaminated soil (PM) and 1.6% in treated soil. CEC and SOC also decreased from about 19 cmol/kg and 0.12% in NF to substantially lower values under ASGM conditions, approximately 6 cmol/kg and 0.06%.
 | Figure 1: Comparison of: (a) Soil pH (H2O), pH (KCl); (b) soil water content (%); (c) cation exchange capacity (cation exchange capacity in cmol/kg); (d) soil organic carbon (%) before and after inoculated with or without plant growth-promoting rhizobacteria+Arbuscular mycorrhizal fungi grown under greenhouse conditions for 6 weeks. Values represent means ± standard error (n = 3). Different letters above the bars indicate statistically significant differences among treatments (P < 0.05). NF: Natural forest, PM: Post mining soil, C: Soil planted with Capsicum frutescens L., M+P: Soil planted with Capsicum frutescens L. inoculated with consortium arbuscular mycorrhizal fungi+plant growth-promoting rhizobacteria. [Click here to view] |
 | Figure 2: Soil N concentration (a), soil K concentration (b), and soil P2O5 concentration (c) with or without plant growth-promoting rhizobacteria+Arbuscular mycorrhizal fungi grown under greenhouse conditions for 6 weeks. Values represent means ± standard error (n = 3). Different letters above the bars indicate statistically significant differences among treatments (P < 0.05). NF: Natural forest, PM: Post mining soil, C: Soil planted with chili, M+P: Soil planted with chili inoculated with consortium arbuscular mycorrhizal fungi+plant growth-promoting rhizobacteria. [Click here to view] |
A similar pattern was observed for total N and available P [Figure 2], both of which were lower under ASGM conditions, whereas total K remained comparatively high. Total N declined from 0.15% in NF to 0.02–0.05% in contaminated soil, and available P declined from 3.4 mg/kg to 1.9–2.9 mg/kg. In contrast, exchangeable K increased from approximately 1.8 cmol/kg to 4.1 cmol/kg in PM, even though it declined by approximately 1.7 cmol/kg following inoculation with the consortium AMF+PGPR. This contrast result suggests that mining disturbance reduced the soil’s buffering capacity, organic matter status, and nutrient-retention potential, while total K may have been less responsive because it is more strongly associated with primary minerals, such as feldspar and mica. This trend is consistent with previous observations in ASGM-impacted soils that reported low SOC (0.95–1.75%), reduced CEC, acidic pH (4.25–5.53), and decreased N and P availability after mining disturbances in Sumbawa, Indonesia [27]. A separate study confirmed that ASGM degrades soil biophysical and chemical properties [7]. In contrast to most nutrients, total K remained high in ASGM soil, plausibly because total K is strongly controlled by primary minerals (e.g., feldspar and mica) and may not decrease rapidly even when available K dynamics change [28].
3.2. Effects of Planting and AMF–PGPR Inoculation On Soil Chemistry and Plant Biomass
Planting C. frutescens, particularly in combination with the AMF+PGPR consortium, was associated with partial improvement in selected soil properties, especially soil pH [Figure 1]. In the post-ASGM medium (PM), soil pH was approximately 3.1 in H2O and 2.8 in KCl, whereas planting increased it to around 3.5 and 3.4, respectively, in the uninoculated treatment (C), and to about 3.2 and 3.3 in the inoculated treatment (M+P). However, this pH improvement was not accompanied by recovery of broader soil quality indicators. Water content decreased from approximately 4.4% in PM to 1.7% in C and 1.4% in M+P, while SOC declined from around 0.06% in PM to 0.03% and 0.02%, respectively [Figure 1]. Total N showed a similar pattern, decreasing from about 0.06% in PM to 0.02% in C and 0.03% in M+P [Figure 2a], indicating that the 6-week planting period was insufficient to rebuild soil organic matter and nitrogen pools. Exchangeable K remained high in PM and C, at approximately 4.1 cmol/kg and 3.7 cmol/kg, respectively, but decreased to about 1.7 cmol/kg in M+P, which was close to the NF value of around 1.8 cmol/kg [Figure 2b]. Available P2O5 was also lowest in M+P, at about 1.9 mg/kg, compared with 2.2 mg/kg in PM and 2.8 mg/kg in C [Figure 2c]. These lower nutrient values in the inoculated treatment may reflect greater nutrient uptake and rhizosphere demand under more active plant–microbe interaction rather than simple nutrient depletion alone. This interpretation is supported by the significantly greater shoot and root dry weights of inoculated seedlings, together with AM colonization in roots [Figure 3], suggesting that AMF+PGPR promoted early plant growth despite persistent soil constraints. These traits are relevant to plant performance in contaminated soil. Phosphate solubilization may improve P availability under stressed soil conditions, whereas nitrogen-fixing ability may support plant growth in nutrient-limited substrates. In combination with AMF inoculation, these bacterial functions may enhance rhizosphere activity and plant establishment during phytoremediation. Synergistic interaction of AMF and PGPR can enhance plant nutrient-use efficiency, nutrient acquisition, and overall plant growth [29]. Overall, these findings indicate partial and short-term improvement in selected soil properties and plant performance, rather than full restoration of ASGM-degraded soil.
 | Figure 3: Mycorrhizal colonization (a), shoot dry weight and root dry weight of Capsicum frutescens L. (b), with or without plant growth-promoting rhizobacteria+Arbuscular mycorrhizal fungi grown under greenhouse conditions for 6 weeks. Values represent means ± standard error (n = 3). Different letters above the bars indicate statistically significant differences among treatments (P < 0.05). NF: Natural forest, PM: Post mining soil, C: Soil planted with Capsicum frutescens L., M+P: Soil planted with Capsicum frutescens L. inoculated with consortium arbuscular mycorrhizal fungi+Plant growth-promoting rhizobacteria. [Click here to view] |
3.3. Multi-Metal Enrichment of Hg, Fe, As, and Al in ASGM-Affected Soil
The ASGM-affected soil showed clear multi-metal enrichment, particularly for total Hg, Fe, As, and Al [Figures 4a, 5a, 6a, and 7a). Compared with the non-contaminated reference (NF), the post-ASGM medium (PM) exhibited a sharp increase in Hg from about 1–2 mg/kg to approximately 44 mg/kg. The amalgamation process directly affects the high Hg concentration in ASGM, which introduces Hg into the processing environment. Fe also increased from around 4,100 mg/kg in NF to 5,500 mg/kg in PM, while As rose from approximately 10 mg/kg to 36–37 mg/kg, and Al from approximately 22,000 mg/kg to 28,000 mg/kg. These results indicate that the studied soil was affected by complex multi-metal contamination rather than Hg enrichment alone. Such a pattern is consistent with ASGM activities, where amalgamation without advance sulfide treatment, due to their technological limitations, including coarse grinding of sulfide-bearing ores to increase gold liberation, contributes directly to Hg inputs. Elevated Al may additionally reflect mineral matrix disturbance and redistribution of fine tailing particles during ore processing and waste deposition. Ore crushing and oxidation of sulfide-bearing minerals indirectly increase the release of As from arsenopyrite and Fe from pyrite [30]. Such impacts are consistent with artisanal gold mining and ore-processing activities that can degrade environmental quality and increase contaminant loading in surrounding systems [31]. Oxidation of sulfide during milling can mobilize As(V) and Fe into the soil/water phases, which may also correlate with Hg retention on Fe-Al oxides [29]. Therefore, the absence of advanced sulfide treatment (e.g., bio-oxidation or flotation) can exacerbate multi-metal contamination because ores are directly ground and mixed with Hg, leading to significant increases in metal (e.g., Hg, Fe, As, and Al) loading through erosion and waste deposition [32].
 | Figure 4: Soil mercury (Hg) concentration (a), shoot and root Hg concentration of Capsicum frutescens L. (b), with or without plant growth-promoting rhizobacteria+Arbuscular mycorrhizal fungi grown under greenhouse conditions for 6 weeks. Values represent means ± standard error (n = 3). Different letters above the bars indicate statistically significant differences among treatments (P < 0.05). NF: Natural forest, PM: Post mining soil, C: Soil planted with Capsicum frutescens L., M+P: Soil planted with Capsicum frutescens L. inoculated with consortium arbuscular mycorrhizal fungi+plant growth-promoting rhizobacteria. [Click here to view] |
 | Figure 5: Soil iron (Fe) concentration (a), shoot and root Fe concentration of Capsicum frutescens L. (b), with or without plant growth-promoting rhizobacteria+Arbuscular mycorrhizal fungi grown under greenhouse conditions for 6 weeks. Values represent means ± standard error (n = 3). Different letters above the bars indicate statistically significant differences among treatments (P < 0.05). NF: Natural forest, PM: Post mining soil, C: Soil planted with Capsicum frutescens L., M+P: Soil planted with Capsicum frutescens L. inoculated with consortium arbuscular mycorrhizal fungi+plant growth-promoting rhizobacteria. [Click here to view] |
 | Figure 6: Soil arsenic (As) concentration (a), shoot and root As concentration of Capsicum frutescens L. (b), with or without plant growth-promoting rhizobacteria+Arbuscular mycorrhizal fungi grown under greenhouse conditions for 6 weeks. Values represent means ± standard error (n = 3). Different letters above the bars indicate statistically significant differences among treatments (P < 0.05). NF: Natural forest, PM: Post mining soil, C: Soil planted with Capsicum frutescens L., M+P: Soil planted with Capsicum frutescens L. inoculated with consortium arbuscular mycorrhizal fungi+Plant growth-promoting rhizobacteria. [Click here to view] |
 | Figure 7: Soil aluminum concentration (a) and soil sulfate concentration (b), with or without plant growth-promoting rhizobacteria+Arbuscular mycorrhizal fungi with Capsicum frutescens L. as biological indicator, grown under greenhouse conditions for 6 weeks. Values represent means ± standard error (n = 3). Different letters above the bars indicate statistically significant differences among treatments (P < 0.05). NF: Natural forest, PM: Post mining soil, C: Soil planted with Capsicum frutescens L., M+P: Soil planted with Capsicum frutescens L. inoculated with consortium arbuscular mycorrhizal fungi+Plant growth-promoting rhizobacteria. [Click here to view] |
Because the soil was steam-sterilized before the experiment, caution is needed in interpreting metal dynamics. Thermal treatment may have altered redox-sensitive phases, sulfur-associated minerals, and the apparent bioavailability of some metals. In particular, the behavior of mercury and arsenic may depend strongly on speciation, which was not determined in this study. Therefore, the observed changes should be interpreted as treatment-associated concentration patterns rather than direct evidence of specific geochemical transformation pathways.
3.4. Effects of Planting on Soil Metal Concentrations: Decreases in Fe, Cd, As, and Al
Following planting, soil metal responses were selective rather than uniform [Figures 4a, 5a, 6a, 7a, and 8a]. Unlike Hg, which remained high and did not differ significantly among PM, C, and M+P (approximately 44–46 mg/kg; [Figure 4a]), several other metals decreased markedly after planting. Soil Fe declined from about 5,500 mg/kg in PM to approximately 300 mg/kg in C and 400 mg/kg in M+P [Figure 5a]. A similar pattern was observed for Cd, which decreased from around 1.55 mg/kg in PM to 0.25 mg/kg in C and 0.30 mg/kg in M+P [Figure 8a]. Soil As also dropped substantially from approximately 36–37 mg/kg in PM to 5–6 mg/kg in both planted treatments [Figure 6a], while Al decreased from about 28,000 mg/kg in PM to roughly 11,000 mg/kg in C and 7,000 mg/kg in M+P [Figure 7a].
 | Figure 8: Soil cadmium (Cd) concentration (a), shoot and root Cd concentration of Capsicum frutescens L. (b), with or without plant growth-promoting rhizobacteria+Arbuscular mycorrhizal fungi grown under greenhouse conditions for 6 weeks. Values represent means ± standard error (n = 3). Different letters above the bars indicate statistically significant differences among treatments (P < 0.05). NF: Natural forest, PM: Post mining soil, C: Soil planted with Capsicum frutescens L., M+P: Soil planted with Capsicum frutescens L. inoculated with consortium arbuscular mycorrhizal fungi+Plant growth-promoting rhizobacteria. [Click here to view] |
This pattern is consistent with possible phytoextraction and/or phytostabilization processes, involving root uptake for some metals and rhizosphere or root-zone immobilization for others, although the reductions were clearly metal-specific and do not indicate complete remediation of the contaminated soil. C. frutescens likely possesses a degree of tolerance to metal stress, and plant performance may improve under AMF colonization (up to 60%), which may reduce metal bioavailability through glomalin production and extraradical hyphal retention [33]. Interestingly, the AMF+PGPR consortium resulted in significantly lower soil Al concentrations than the uninoculated treatment. This may reflect the synergy between (i) Al adsorption/retention by AMF hyphae, (ii) PGPR production of Fe/Al-binding siderophores, and (iii) improved soil aggregation that reduced erosion-driven redistribution of ASGM waste. Such combined effects can be stronger in multi-metal contaminated soils and are broadly consistent with substantial Cd reduction and improved biomass reported under comparable contaminated conditions [34].
Although soil Fe, Cd, As, and Al concentrations decreased after planting, the present dataset does not permit definitive mass-balance attribution of these reductions. The observed declines may reflect a combination of plant uptake, rhizosphere immobilization, redistribution within the pot soil, and heterogeneity in contaminated substrate. Because leachate, sequential extraction, and metal speciation were not analyzed, the fate of each metal should be interpreted cautiously.
3.5. Increased Soil Sulfate Under Inoculation: Tentative Rhizosphere Sulfur Cycling Interpretation
In contrast to soil Al, soil sulfate was highest in inoculated treatments [Figure 7b]. Sulfate concentration reached approximately 870 mg/kg in M+P, compared with around 480 mg/kg in the uninoculated planted treatment (C), and about 730–740 mg/kg in NF and PM. Thus, planting alone was associated with a decrease in soil sulfate, whereas AMF+PGPR inoculation appeared to reverse this pattern. This response may be linked to enhanced rhizosphere-driven sulfur cycling, particularly the oxidation/mineralization of organic S to inorganic sulfate (SO42−) mediated by plant–microbe interactions. Although root biomass, root exudation, and sulfur-transforming functional groups were not quantified in this study, AMF+PGPR inoculation has been reported to stimulate rhizosphere activity and nutrient turnover, which could plausibly accelerate S mineralization. Although the elevated SO42− in inoculated soils is interpreted as an indicator of intensified S transformation, this mechanistic explanation remains tentative in the absence of sulfur speciation and functional microbiological evidence and should be verified in future work.
From an agronomic perspective, increased SO42− may improve short-term S availability for plants. Nevertheless, sulfate accumulation is not inherently beneficial and may entail environmental trade-offs. Under conditions favoring solute transport – such as coarse-textured soils, high rainfall/irrigation regimes, or repeated S inputs – elevated SO42− can increase leaching potential to groundwater. Higher soluble ion loads may also raise soil electrical conductivity and influence nutrient balance. Given the greenhouse pot setting and limited experimental duration, this study did not assess sulfate retention, leaching losses, or off-site impacts. Therefore, longer-term and field-scale assessments are needed, including SO42− mass balance (inputs–plant uptake–outputs), leachate/soil pore-water measurements, EC dynamics, and crop uptake, to determine whether inoculation-driven S transformation provides sustained agronomic benefits without adverse environmental consequences.
3.6. Differential Accumulation of Heavy Metals in Plant Tissues and Underlying Rhizospheric Mechanisms
The present study showed that microbial inoculation produced metal-specific patterns of accumulation in plant tissues rather than a uniform decrease across all heavy metals. Under AMF+PGPR treatment, Hg, Zn, and As concentrations were reduced in both roots and shoots relative to uninoculated seedlings [Figures 4b, 6b, and 9b]. For example, root Hg decreased from approximately 9.8 mg/kg in C to about 0.1 mg/kg in M+P, while shoot Hg declined from around 0.35 mg/kg to 0.20 mg/kg. A similar trend was observed for As, with root concentrations decreasing from approximately 0.97 mg/kg in C to 0.18 mg/kg in M+P, and shoot concentrations from 0.54 mg/kg to 0.37 mg/kg. Root Zn also declined sharply from about 185 mg/kg in C to roughly 7–8 mg/kg in M+P, although shoot Zn remained similar at around 103–104 mg/kg in both treatments. At the soil level, Zn concentration was higher in planted treatments, especially under inoculation, as shown in Figures 9a.
 | Figure 9: Soil zinc (Zn) concentration (a), shoot and root Zn concentration of Capsicum frutescens L. (b), with or without plant growth-promoting rhizobacteria+Arbuscular mycorrhizal fungi grown under greenhouse conditions for 6 weeks. Values represent means ± standard error (n = 3). Different letters above the bars indicate statistically significant differences among treatments (P < 0.05). NF: Natural forest, PM: Post mining soil, C: Soil planted with Capsicum frutescens L., M+P: Soil planted with Capsicum frutescens L. inoculated with consortium arbuscular mycorrhizal fungi+Plant growth-promoting rhizobacteria. [Click here to view] |
Soil Mn was undetectable in NF and PM but became measurable in planted treatments, with the highest concentration observed in M+P [Figure 10a]. In contrast, Fe and Mn showed a different pattern: Shoot concentrations were maintained or increased under inoculation, whereas root concentrations declined strongly [Figures 5b and 10b]. Shoot Fe increased from approximately 0.39 mg/kg in C to 0.77 mg/kg in M+P, while root Fe decreased from about 175 mg/kg to only ~4 mg/kg. Similarly, shoot Mn increased from around 0.62 mg/kg in C to 1.29 mg/kg in M+P, whereas root Mn declined from approximately 0.63 mg/kg to nearly undetectable levels. Cd showed another distinct response: Root Cd decreased from about 1.68 mg/kg in C to nearly 0 mg/kg in M+P, yet shoot Cd increased from approximately 0.68 mg/kg to 0.92 mg/kg [Figure 8b]. Together, these results indicate selective regulation of metal uptake and translocation rather than uniform detoxification across all heavy metals.
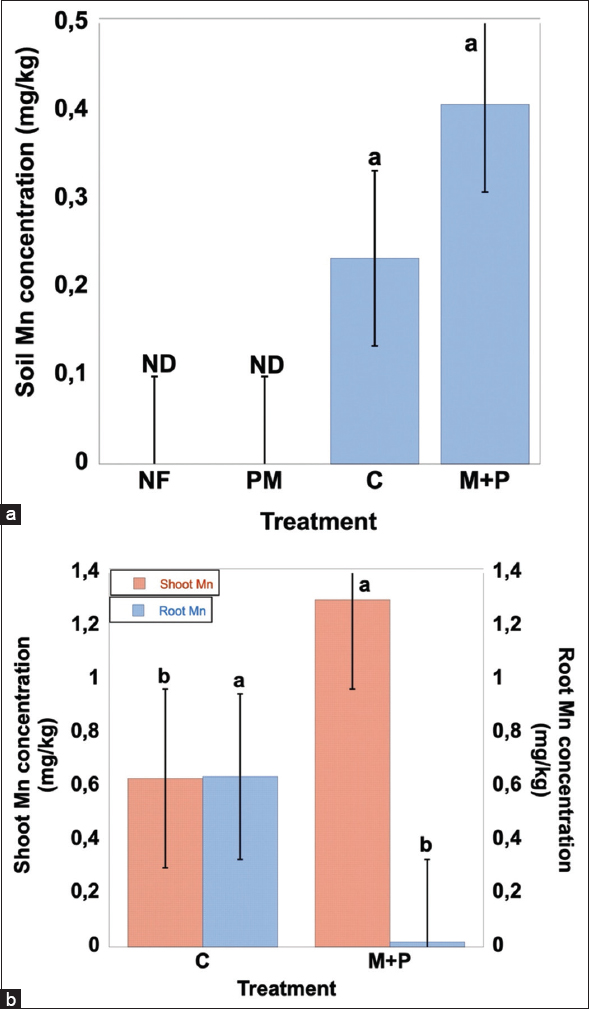 | Figure 10: Soil manganese (Mn) concentration (a), shoot and root Mn concentration of Capsicum frutescens L. (b), with or without Plant growth-promoting rhizobacteria+Arbuscular mycorrhizal fungi grown under greenhouse conditions for 6 weeks. Values represent means ± standard error (n = 3). Different letters above the bars indicate statistically significant differences among treatments (P < 0.05). NF: Natural forest, PM: Post mining soil, C: Soil planted with Capsicum frutescens L., M+P: Soil planted with Capsicum frutescens L. inoculated with consortium arbuscular mycorrhizal fungi+Plant growth-promoting rhizobacteria. [Click here to view] |
The reduction in Hg, As, and root Zn under AMF+PGPR treatment is consistent with rhizospheric immobilization and fungal sequestration. AMF are known to retain toxic metals within hyphae and vesicles and to bind metals through glomalin-related soil proteins, while PGPR may contribute through extracellular polymeric substances, biosorption, siderophore-mediated interactions, or other microbially mediated transformations that reduce metal bioavailability. For highly toxic and non-essential elements such as Hg and As, these mechanisms may help limit internal transport and reduce tissue accumulation. The combined action of AMF-derived glomalin, PGPR activity, and rhizosphere pH modification may therefore contribute to reduced bioavailability and support biomass maintenance under high contamination [37].
By contrast, the continued or enhanced translocation of Fe and Mn to shoots suggests that inoculation did not simply suppress metal movement, but may have selectively favored the transport of physiologically important micronutrients under stress. Fe and Mn are essential elements for photosynthesis, redox regulation, and enzymatic activity, and their greater shoot accumulation in M+P may reflect improved nutrient acquisition rather than toxic overload. The Cd response was more complex because inoculation strongly reduced root accumulation but did not suppress shoot transport. This suggests that AMF+PGPR effects were element-specific and involved differential regulation of root retention, mobilization, and translocation. Overall, these findings support selective regulation of metal uptake and partitioning rather than a generalized detoxification mechanism.
3.7. Elevation of Shoot Fe and Mn: Nutrition Improvement and Adaptation Effects
In contrast to Hg, Zn, and As, the AMF+PGPR consortium was associated with greater translocation of Fe and Mn to shoots [Figures 5b and 10b]. Shoot Fe increased from approximately 0.39 mg/kg in C to 0.77 mg/kg in M+P, while shoot Mn increased from around 0.62 mg/kg to 1.29 mg/kg. At the same time, root Fe declined sharply from about 175 mg/kg in C to ~4 mg/kg in M+P, and root Mn decreased from approximately 0.63 mg/kg to nearly undetectable levels. This pattern suggests altered partitioning of Fe and Mn under inoculation, with reduced retention in roots and greater allocation to shoots.
Unlike Hg, As, and Zn (in excess), Fe and Mn are essential micronutrients involved in chlorophyll synthesis, photosynthetic electron transport, and antioxidant enzyme systems, including superoxide dismutase and peroxidases. Under heavy metal stress, plants often require increased antioxidant capacity, thereby elevating the demand for Fe and Mn. PGPR-mediated siderophore production can effectively enhance Fe solubilization and uptake, while AMF hyphal networks expand the effective absorption area of roots. Under heavy metal stress, plants often increase antioxidant demand, potentially increasing Fe and Mn requirements. PGPR siderophores may enhance Fe solubilization and uptake, while AMF hyphal networks expand the effective absorption zone. Therefore, the increased Fe and Mn in shoots likely reflect improved micronutrient allocation and stress adaptation rather than non-selective metal uptake [38]. These findings further support selective regulation of metal translocation among elements, rather than a uniform reduction in metal mobility.
3.8. Cd Behavior: Partial Regulation without Complete Restriction of Translocation
Cd exhibited a distinct pattern compared with the other metals. Although root Cd concentrations were substantially reduced under inoculation, shoot Cd levels were not significantly suppressed [Figure 8b]. Root Cd declined from approximately 1.68 mg/kg in C to nearly undetectable levels in M+P, whereas shoot Cd increased from about 0.68 mg/kg to 0.92 mg/kg. This indicates that, despite the marked reduction in root Cd accumulation, Cd was still translocated to shoots through the xylem. PGPR has been reported to modulate the expression of metal transporters in roots, but does not completely block Cd translocation under multi-metal conditions [39]. In addition, uninoculated seedlings had lower shoot uptake because growth stress restricted overall translocation, whereas inoculation increased biomass and may thereby have increased metal flux to the shoots [35]. Furthermore, Cd shares transport pathways with Zn and calcium due to ionic similarity and can enter roots through non-specific divalent cation transporters. Its relatively high mobility within the xylem may explain continued shoot translocation despite reduced root concentrations. Moreover, AMF-mediated sequestration efficiency for Cd has been reported to vary depending on fungal species and soil chemistry. The observed pattern, therefore, suggests partial rhizospheric regulation without complete restriction of long-distance Cd transport [40-42].
Taken together, these results indicate that microbial inoculation influences metal dynamics in a metal-specific manner, shaped by chemical properties, plant nutritional requirements, and rhizosphere interactions. Shoots require critical micronutrients (Fe, Mn) for metabolic and antioxidative processes, while highly toxic, non-essential metals such as Hg are more readily removed. Meanwhile, Cadmium has intermediate behavior due to chemical mimicry and transport mechanisms. This targeted regulation highlights the intricacy of plant-microbe-metal interactions in polluted soils. Overall, these results indicate selective regulation of metal uptake and translocation rather than uniform detoxification of all heavy metals.
3.9. Rhizosphere Viable Bacterial Abundance Supports Consortium Establishment (CFU)
As shown in Table 1, plate count enumeration conducted at week 6 revealed a significant disparity in viable bacterial abundance across treatments. The control showed a minimal bacterial count (5 CFU/mL in soil suspension), whereas the consortium treatment (AMF+PGPR) showed a much higher abundance (4.5 × 104 CFU/mL in soil suspension). The growing media was steam-sterilized (100°C for 5 h) and verified as sterile using agar plating (no visible colonies) before planting. This result indicates that the bacterial populations observed at week 6 represent microbial establishment or recolonization during the cultivation period. The endpoint CFU findings demonstrate that the consortium application was related to a significantly increased viable bacterial population in the rhizosphere by week 6. The higher CFU count in the consortium treatment suggests that viable bacteria persisted in the rhizosphere following inoculation. The near absence of bacteria in the control indicates that rhizosphere recolonization under non-aseptic pot conditions was minimal without inoculation; however, potential microbial introduction from irrigation water, airborne deposition, or seedling-associated microbiota during the 6-week experiment cannot be excluded.
Table 1: Colony number of bacteria in soil with or without inoculation of AMF+PGPR in comparison to the initial soil and the control soil 6 weeks after treatment.
| No. | Treatment | Colony number (CFU/mL) |
|---|---|---|
| 1. | Initial soil (Sterilized) | 0 b |
| 2. | Control soil | 5 b |
| 3. | Soil inoculated AMF+PGPR | 4.5×104 a |
Values are presented as mean±standard error (n=3). Different letters indicate statistically significant differences among treatments according to Tukey’s honest significant difference post hoc test at P<0.05. AMF: Arbuscular mycorrhizal fungi, PGPR: Plant growth-promoting rhizobacteria, CFU: Colony-forming unit.
The findings strengthen the theory that the AMF+PGPR treatment likely facilitated the observed rhizosphere microbial enrichment, which may partly explain the enhanced early plant performance reported in this study through mechanisms typically associated with PGPR, such as enhanced nutrient acquisition, modulation of stress responses, and improved root-microbe interactions. Nevertheless, because bacterial enumeration was performed solely at week 6, the data affirm endpoint viability but do not elucidate week-to-week persistence dynamics. Subsequent research should integrate time-series rhizosphere monitoring and, when possible, strain-level verification to differentiate implanted PGPR from recolonizing background microbiota.
3.10. Plant Survival Dynamics: Transient Early Benefits without Improved Final Survival
Plant survival decreased markedly in both treatments, particularly during the first 2 weeks [Figure 11]. In the control (SR-C), survival declined from 100% at week 0 to 27% at week 2, then decreased further to 18% at week 4 and remained at 18% at week 6. Under AMF+PGPR inoculation (SR-AMF+PGPR), survival was initially higher, declining from 100% at week 0 to 55% at week 2 and 45% at week 4, but then fell to 19% by week 6, essentially converging with the control. These results indicate that AMF+PGPR improved early survival during establishment (weeks 2–4) but did not improve final survival at the end of the experiment.
.jpg) | Figure 11: Survival rate (%) of Capsicum frutescens in artisanal and small-scale gold mining-affected soil with and without plant growth-promoting rhizobacteria+ Arbuscular mycorrhizal fungi inoculation over 6 weeks (n = 21 plants per treatment). [Click here to view] |
The sharp decline during weeks 0–2 identifies a critical stress phase in ASGM-affected soil, during which inoculation appeared to provide a temporary buffering effect on plant establishment. However, the convergence of survival by week 6 suggests that major constraints persisted beyond the protective capacity of the inoculum. These likely included residual phytotoxicity from bioavailable metal fractions, nutrient limitations or imbalances, and adverse soil physical conditions that hindered root development and disrupted water–air balance. Thus, despite improvements in several soil indicators, the very low final plant survival indicates that these changes were insufficient to support full biological recovery within the experimental period. Accordingly, the remediation potential of AMF+PGPR inoculation in this context should be regarded as partial and temporally constrained.
This interpretation is further supported by the relatively large error bars observed across several parameters, suggesting substantial variability among replicates. Such variability may partly reflect the limited number of analytical replicates (n = 3), the heterogeneous nature of ASGM-impacted soil, and biological variation among seedlings, including possible differences in AMF and PGPR establishment. Nevertheless, several parameters remained significantly different across treatments, indicating that some treatment effects remained detectable despite the observed heterogeneity. Therefore, while the trends remain informative, particularly in indicating partial improvement and element-specific responses, they should be interpreted cautiously and not as evidence of uniformly consistent treatment effects.
Comparison with previous microbial-assisted phytoremediation studies suggests that the present findings follow a pattern commonly reported in contaminated soils: microbial inoculation may improve selected soil properties and modify plant–contaminant interactions, but its effectiveness can be limited under severe contamination stress [44,46]. PGPR–AMF inoculation has been reported to improve plant performance in petroleum-contaminated saline-alkali soil [43], while PGPR-based systems and AMF-based systems have separately been shown to alleviate heavy-metal stress and improve plant performance under contaminated or stress conditions [45,46]. However, under severe metal stress, these benefits may remain incomplete because excessive contamination can inhibit root growth, reduce microbial colonization efficiency, and constrain plant survival [46]. Taken together, the present study demonstrates partial soil improvement and selective modification of metal responses following planting and AMF+PGPR inoculation, but not full restoration of ASGM-degraded soil.
4. CONCLUSION
This study shows that ASGM substantially degraded soil quality and generated multi-metal contamination, as reflected by reduced soil fertility and elevated concentrations of Hg, Fe, As, and Al. Establishment of C. frutescens in contaminated soil was associated with reductions in several soil metal concentrations, while inoculation with the AMF+PGPR consortium contributed to higher soil pH, greater root colonization, and significantly increased plant biomass, indicating improved early plant establishment under stressed conditions.
Importantly, the effect of microbial inoculation on metal accumulation was metal-specific rather than uniform. The AMF+PGPR consortium reduced the accumulation of highly hazardous metals, including Hg, As, and Zn, in plant tissues, consistent with rhizospheric immobilization and fungal sequestration. In contrast, translocation of Fe and Mn to shoots was maintained or enhanced, suggesting continued micronutrient transport under stress, whereas Cd showed partial limitation of root accumulation but remained detectable in shoots.
Overall, the AMF+PGPR consortium supported partial and early-stage remediation-related responses in ASGM-contaminated soil, but these effects were metal-specific and not sufficient to indicate full restoration. Therefore, this approach should be interpreted as a potentially useful but still limited biological strategy, and further long-term and field-scale studies are needed to determine whether these benefits can be sustained under highly contaminated conditions.
5. ACKNOWLEDGMENTS
The authors gratefully acknowledge the financial support provided by the Directorate General of Higher Education, Ministry of Education, Culture, Research, and Technology of Indonesia under Contract No. 050/DirDPPM/70/DPPM/PTM-KEMDIKTISAINTEK/VI/2025 for the 2025 fiscal year.
6. AUTHORS’ CONTRIBUTIONS
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agreed to be accountable for all aspects of the work. All the authors are eligible to be author as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
7. CONFLICT OF INTEREST
The authors report no financial or any other conflicts of interest in this work.
8. ETHICAL APPROVALS
This study does not involve experiments on animals or human subjects.
9. DATA AVAILABILITY
The data supporting the findings of this study are not publicly available but are available from the corresponding author upon reasonable request.
10. PUBLISHER’S NOTE
All claims expressed in this article are solely those of the authors and do not necessarily represent those of the publisher, the editors and the reviewers. This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
11. USE OF ARTIFICIAL INTELLIGENCE (AI)-ASSISTED TECHNOLOGY
The authors declare that they have not used artificial intelligence (AI)-tools for writing and editing of the manuscript, and no images were manipulated using AI.
REFERENCES
1. Suhartini S, Abubakar. Socio-economic impacts and policy of artisanal small-scale gold mining in relation to sustainable agriculture:A case study at Sekotong of West Lombok. J Degraded Min Lands Manag. 2017;4(3):789-96. [CrossRef]
2. Krisnayanti BD, Vassura I, Asmara MD, Ekawanti A, Suheri H. Analysis of artisanal small-scale gold mining sector in West Sumbawa Regency, Indonesia. J Health Pollut. 2016;6(12):26-33. [CrossRef]
3. Craw D, Palmer M. Anthropogenic Hg-Au residues from placer mining compared to natural Au-Hg sources, South Island, New Zealand. N Z J Geol Geophys. 2025;68(2):342-61. [CrossRef]
4. Owusu-Prempeh N, Awuah OK, Abebrese IK, Amaning EN. Analysis of the status and ecological risks of heavy metals contamination in artisanal and small-scale gold mine-spoils at the Atewa forest landscape, Ghana. Sci Afr. 2022;16:e01235. [CrossRef]
5. Tun AZ, Wongsasuluk P, Siriwong W. Heavy metals in the soils of placer small-scale gold mining sites in Myanmar. J Health Pollut. 2020;10(27):200911. [CrossRef]
6. Quiceno-Vallejo MF, Escobar MC, Vásquez Y. Impact of mine drainage on the microbial community of the soil. Rev Acad Colomb Cienc Ex Fis Nat. 2020;44(170):241-56. [CrossRef]
7. Asare D, Boateng K, Yeboah-Assiamah E. Impact of different illegal artisanal small-scale mining methods on soil physical, chemical and microbial properties four years after mining. Environ Chall. 2024;100:103318 [CrossRef]
8. Yuwono SB, Banuwa IS, Suryono N, Somura H, Dermiyati D. Mercury pollution in the soil and river water of the Ratai watershed by artisanal and small-scale gold mining activities in Pesawaran District, Lampung, Indonesia. J Degraded Min Lands Manag. 2023;10(2):4233-43. [CrossRef]
9. Tuheteru EJ, Tuheteru FD, Hartami PN, Burhannudinnur M, Prakoso S, Husna H, et al. Soil properties change, and Arbuscular mycorrhizal fungi associated with plants growing on the post-gold mining land of Bombana, Indonesia. J Degraded Min Lands Manag. 2023;11(1):4863-73. [CrossRef]
10. Lugendo IJ, Mwesiga V. Heavy metal contamination and health risks in Katoro's artisanal gold mining soils, Tanzania. Tanzania J Sci. 2025;51(3):774. [CrossRef]
11. Zheng X, Cao H, Liu B, Zhang M, Zhang C, Chen P, et al. Effects of mercury contamination on microbial diversity of different kinds of soil. Microorganisms. 2022;10(5):977. [CrossRef]
12. Xiao R, Wang S, Li R, Wang JJ, Zhang Z. Soil heavy metal contamination and health risks associated with artisanal gold mining in Tongguan, Shaanxi, China. Ecotoxicol Environ Saf. 2017;141:17-24. [CrossRef]
13. Scammacca O. Mining Risk Assessment at the Territory Scale:Development of a Tool Tested on the Example of Gold Mining in French Guiana. HAL (le Centre Pour la Communication Scientifique Directe);2020. Available from: https://hal.archives-ouvertes.fr/tel-03225840 [Last accessed on 2026 Jan 28].
14. Kumar V, Umesh M, Shanmugam MK, Chakraborty P, Duhan L, Gummadi SN, et al. A retrospection on mercury contamination, bioaccumulation, and toxicity in diverse environments:Current insights and future prospects. Sustainability. 2023;15(18):13292. [CrossRef]
15. Tabelin CB, Silwamba M, Paglinawan FC, Mondejar AJ, Duc HG, Resabal VJ, et al. Solid-phase partitioning and release-retention mechanisms of copper, lead, zinc and arsenic in soils impacted by artisanal and small-scale gold mining (ASGM) activities. Chemosphere. 2020;260:127574. [CrossRef]
16. Akansha J, Thakur S, Sai Chaithanya M, Sen Gupta B, Das S, Das B, et al. Technological and economic analysis of electrokinetic remediation of contaminated soil:A global perspective and its application in Indian scenario. Heliyon. 2024;10(2):e24293. [CrossRef]
17. U.S. Environmental Protection Agency. Remediation Case Studies:Thermal Desorption, Soil Washing, and in Situ Vitrification. United States:Environmental Protection Agency;1995.
18. Qin H, Wang Z, Sha W, Song S, Qin F, Zhang W. Role of plant-growth-promoting rhizobacteria in plant machinery for soil heavy metal detoxification. Microorganisms. 2024;12(4):700. [CrossRef]
19. Wulandari D, Saridi, Cheng W, Tawaraya K. Arbuscular mycorrhizal fungal inoculation improves Albizia saman and Paraserianthes falcataria growth in post-opencast coal mine field in East Kalimantan, Indonesia. Forest Ecol Manag. 2016;376:67-73. [CrossRef]
20. Zhao S, Lin Q, Wang J, Chen Y. Arbuscular mycorrhizal Fungi-assisted phytoremediation. Microorganisms. 2024;12(5):1034.
21. Zhang X, Li Y, Ma W, Su Q, Zhang H. Sustainable remediation of soil and water utilizing Arbuscular mycorrhizal fungi:Mechanisms, recent advances and future perspectives. Microorganisms. 2024;12(4):910. [CrossRef]
22. Wulandari D, Nufus M, Faridah E, Maulana AF, Tawaraya K. Native Arbuscular mycorrhizal fungi and Nauclea orientalis for potential reclamation of tropical coal mining areas. Environ Adv. 2024;15:100462. [CrossRef]
23. Giovanetti M, Mosse B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 1980;84(3):489-500. [CrossRef]
24. U.S. EPA. Method 3050B:Acid Digestion of Sediments, Sludges, and Soils (Revision 2). U.S. Environmental Protection Agency;1996. Available from: https://settek.com/wp-content/uploads/2023/11/epa-method-3050b.pdf [Last accessed on 2026 Jan 30].
25. Agus F, Subiksa IG. Peatlands:Potential for Agriculture and Environmental Aspects. Bogor, Indonesia:Soil Research Institute;2008.
26. Murphy J, Riley JP. A modified single solution method for the determination of phosphate in natural waters. Anal Chim Acta. 1962;27:31-6. [CrossRef]
27. Salam RH, Suwardji S, Fauzi T, Kusnarta IG. Characteristics of physical and chemical properties of soil after gold processing (case study of unlicensed gold mining in the District Sumbawa Regency, Indonesia). Int J Sci Technol Manag. 2024;5(3):591-9. [CrossRef]
28. Madaras M, KoubováM. Potassium availability and soil extraction tests in agricultural soils with low exchangeable potassium content. Plant Soil Environ. 2015;61(5):234-9. [CrossRef]
29. Zhang F, Wang Y, Alowaifeer A, Wang S, Li J, Chen J. Solid-phase partitioning and release–retention mechanisms of Cu, Pb, Zn and As in soils impacted by artisanal and small-scale gold mining (ASGM) activities. Sci Total Environ. 2020;736:139583. [CrossRef]
30. Ardha N, Saleh N, Damayanti R. Konsep desain custom plant flotasi untuk mengolah bijih sulfida marjinal mengandung emas/perak. J Teknol Mineral Batubara. 2014;10(1):1-14.
31. Torrance KW, Redwood SD, Cecchi A. The impact of artisanal gold mining, ore processing and mineralization on water quality in Marmato, Colombia. Environ Geochem Health. 2021;43(10):4265-82. [CrossRef]
32. Esdaile LJ, Chalker JM. The mercury problem in artisanal and small-scale gold mining. Chem Rev. 2018;118(2):231-69. [CrossRef]
33. Li L, Shi Y, Xia W, Wang X, Xin Z, Liao Y, et al. Soil amendments altered arbuscular mycorrhizal fungal communities in cadmium-contaminated vegetable fields. Front Microbiol. 2024;15:1470137. [CrossRef]
34. Rasouli-Sadaghiani M, Khodaverdiloo H, Barin M, Kazemalilou S. Influence of PGPR bacteria and arbuscular mycorrhizal fungi on growth and some physiological parameters of Onopordon acanthium in a Cd-contaminated soil. Water Soil. 2016;30(2):542-54.
35. Yu L, Zhang H, Zhang W, Liu K, Liu M, Shao X. Cooperation between arbuscular mycorrhizal fungi and plant growth-promoting bacteria and their effects on plant growth and soil quality. PeerJ. 2022;10:e13080. [CrossRef]
36. Kuila D, Ghosh S. Aspects, problems and utilization of arbuscular mycorrhizal (AM) application as bio-fertilizer in sustainable agriculture. Curr Res Microbiol Sci. 2022;3:100107. [CrossRef]
37. Hnini M, Rabeh K, Oubohssaine M. Interactions between beneficial soil microorganisms (PGPR and AMF) and host plants for environmental restoration:A systematic review. Plant Stress. 2024;11:100391. [CrossRef]
38. Rao MC, Rani AP, Tejaswi MR, Harshitha Y, Kumar NS, Bindu SH, et al. Enhancing the phytoremediation of heavy metals by plant growth promoting rhizobacteria (PGPR) consortium:A narrative review. J Basic Microbiol. 2025;65(4):e2400529. [CrossRef]
39. Khanna K, Jamwal VL, Gandhi SG, Ohri P, Bhardwaj R. Metal resistant PGPR lowered Cd uptake and expression of metal transporter genes with improved growth and photosynthetic pigments in Lycopersicon esculentum under metal toxicity. Sci Rep. 2019;9:5855. [CrossRef]
40. Kuang Y, Li X, Wang Z, Wang X, Wei H, Chen H, et al. Effects of arbuscular mycorrhizal fungi on the growth and root cell ultrastructure of Eucalyptus grandis under cadmium stress. J Fungi (Basel). 2023;9(2):140. [CrossRef]
41. Kanwal S, Bano A, Malik RN. Effects of arbuscular mycorrhizal fungi on metals uptake, physiological and biochemical response of Medicago sativa L. With increasing Zn and Cd concentrations in soil. Am J Plant Sci. 2015;6:2906-23. [CrossRef]
42. Ma C, Wu C, Han H, Bai D, Zhang Z, Li Y, et al. Metabolomics reveals concentration-specific adaptive mechanisms of arbuscular mycorrhizal fungi in cadmium translocation and detoxification in arundinoideae (Phragmites australis). Plant Cell Environ. 2026;49(1):514-30. [CrossRef]
43. Xun F, Xie B, Liu S, Guo C. Effect of plant growth-promoting bacteria (PGPR) and arbuscular mycorrhizal fungi (AMF) inoculation on oats in saline-alkali soil contaminated by petroleum to enhance phytoremediation. Environ Sci Pollut Res Int. 2015;22:598-608. [CrossRef]
44. Ma Y, Ankit, Tiwari J, Bauddh K. Plant-mycorrhizal fungi interactions in phytoremediation of geogenic contaminated soils. Front Microbiol. 2022;13:843415. [CrossRef]
45. Anbuganesan V, Vishnupradeep R, Bruno LB, Sharmila K, Freitas H, Rajkumar M. Combined application of biochar and plant growth-promoting rhizobacteria improves heavy metal and drought stress tolerance in Zea mays. Plants (Basel). 2024;13(8):1143. [CrossRef]
46. Ferrol N, Tamayo E, Vargas P. The heavy metal paradox in arbuscular mycorrhizas:From mechanisms to biotechnological applications. J Exp Bot. 2016;67(22):6253-65. [CrossRef]